
|
By Sheridan Wilkinson, Undergraduate Research Student Donax variabilis (Coquina clams) are small, wedge-shaped clams that dominate the intertidal zone by filter feeding on unicellular algae and detritus. Coquina clams are found along the Gulf Coast and along most of the East Coast of the United States (Ruppert and Fox, 1988). In fact, coquinas have been reported to constitute up to 95% of the macrofaunal biomass of sandy beaches (McLachlan et. al., 1981); however, they use their foot (a fleshy muscle found in molluscs for movement) to burrow into the sand where they often remain unseen (de la Huz et. al., 2002; Nel et. al., 2001). Coquina clams have also been shown to be an important food resource for both sublittoral and supralittoral predators, including surf fish such as Florida pompano (Trachinotus carolinus) (Leber, 1982), stingrays (Jablonski, 2001), Moon snails (Polinices duplicatus) (Tiffany, 1971), and shore birds (Schneider, 1982; Jablonski, 2001). Given their importance in the food web of sandy beach ecosystems, coquinas are considered an indicator species for the ecosystem health, especially given their prevalence and widespread distribution.  Donax spp. are dioecious (separate sexes) (Ansell, 1983) and have spawning events between January and May, which produces a planktonic, veliger larva that inhabits the pelagic zone of the water column (Cobb et. al., 2011). Larval duration has been estimated to be 3—4 weeks (Ansell, 1983) before they undergo metamorphosis into the adult stage (Chanley, 1969), settle to the bottom, and migrate to the swash zone of sandy shores (Jablonski, 2001). During the larval stage, Coquina clams may be subject to ocean currents like the Gulf Stream and Loop Current that could potentially distribute the larvae over great distances along the coast line (Stommel, 1958; Gordon, 1967). Currently, D. variabilis is described as only one species despite the great distance coquina larvae would have to travel (over a 4,828-km geographic range from Texas to New York), varying habitat, and oceanic and geographic barriers to maintain genetic connectivity. 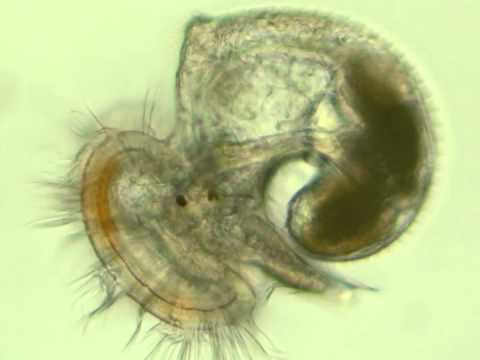 In my study, I am comparing the genetic variation of populations throughout the distribution of D. variabilis using the mitochondrial COI (cytochrome c oxidase I) marker which contributes to the electron transport chain, as well as the mitochondrial 16S rRNA marker which is part of the ribosomal RNA subunit. Both of these mitochondrial markers will be used to help determine if speciation has occurred in D. variabilis throughout the geographic distribution. These roughly 600 base-pair regions are present in nearly every animal due to their importance within cell functions and are, therefore, ideal genes to use in DNA barcoding to deduce the level of speciation between and within species. It is unlikely that throughout the natural distribution of D. variabilis geneflow is present between every population; instead, it is far more plausible that oceanic or geographic barriers and natural selection have led to phylogeographic breaks and structure.
Literature Cited: Ansell, A.D. 1983. Sandy beaches as ecosystems: the biology of the genus Donax. In Developments in Hydrobiology. 19, 608-635 Chanley, P. 1969. Larval Development of the Coquina Clam, Donax Variabilis Say, with a Discussion of the Structure of the Larval Hinge in the Tellinacea. Bulletin of Marine Science, 19.1, 214-224 Cobb, J.C., S.P. Stephenson, W.S. Arnold. 2011. Reproductive cycle and recruitment patterns of the coquina clam Donax variabilis say along the central Gulf coast of Florida. J. Shellfish Res. 30(1), 25-34 de la Huz, R., M. Lastra, J. Lopez. 2002. The influence of sediment grain size on burrowing, growth and metabolism of Donax trunculus L. (Bivalvia: Donacidae). J. of Sea Res. 47(2), 85-95 Gordon, A.L. 1967. Circulation of the Caribbean Sea. J. Geophys. Res. 72, 6207-6223 Jablonski, E. 2001. The population ecology and reproductive biology of coquina clams (Donax variabilis) on a Florida beach. Leber, K.M. 1982. Bivalves (Tellinacea, Donacidae) on a North Carolina beach--contrasting population-size structures and tidal migrations. Marine Ecology Progress Series 7(3), 297-301 McLachlan, A et al. 1981. Sand beach energetics: An ecosystem approach towards a high energy interface. Estuar. Coast. Shelf Sci. 13(1), 11-25 Nel, Ronel, Anton McLachlan, Deo P.E. Winter. 2001. The effect of grain size on the burrowing of two Donax species. J. of Experimental Marine Biology and Ecology. 265(2), 219-238 Ruppert, H., R. Fox. 1988. Seashore Animals of the Southeast. Columbia: University of South Carolina Press. Schneider, D. 1982. Predation by ruddy turnstones (Arenaria interpres) on a polymorphic clam (Donax variabilis) at Sanibel Island, Florida. Bulletin of Marine Science. 32(1), 341-344 Stommel, H. M. 1958. The Gulf Stream: A Physical and Dynamical Description. California: University of California Press. Tiffany, W.J. 1971. The tidal migration of Donax variabilis Say (Mollusca: Bivalvia). Veliger. 14(1), 82-85
2 Comments
|
Janosik LabWelcome to our lab blog. Members of the lab will write posts about current research, science or topics of interest. We look forward to your comments! Archives
December 2017
Categories |
 RSS Feed
RSS Feed